

Tradicionalmente, las cepas de levadura para vinos espumosos elaborados con el método tradicional se han seleccionado por sus caracteres fenotípicos tales como capacidad fermentativa en condiciones restrictivas de pH, presión y limitación de nutrientes (Bartra, 1995). Estas características pueden variar durante su multiplicación (crecimiento vegetativo), debido a cambios genéticos (inestabilidad cariotípica) que puede traducirse en cambios que afectan al fenotipo por el cual la cepa se había seleccionado. Si estos cambios ocurren durante los procesos de producción industrial pueden llegar a comprometer la producción, ya que generalmente se manifiestan demasiado tarde para su remedio. Un caso típico son las paradas fermentativas, pero también podríamos añadir la aparición de aromas inesperados y cambios en la proporción de glicerol y otros metabolitos secundarios. La colaboración entre enólogos y genéticos puede actualmente anticipar estos cambios y predecir el comportamiento industrial de una determinada cepa de levadura.
Las cepas de levadura, tanto industriales como silvestres, pueden presentar altas frecuencias de cambios en su cariotipo durante el crecimiento vegetativo. Estos cambios, a los que designamos como inestabilidad cariotípica, consisten básicamente en reorganizaciones cromosómicas. La elaboración de cavas y otros vinos espumosos de calidad requiere la producción de un pie de cuba o inóculo de levadura adaptado a las características del vino base (pH, grado alcohólico, acidez, dióxido de azufre, etc.). Seguidamente, este inóculo debe multiplicarse en el mismo vino base hasta llegar a una población de 106 levaduras por mL de cuvé o vino base. Este crecimiento puede dar origen a poblaciones distintas de la original inoculada, con características negativas si la estabilidad genética de la cepa no es la adecuada.
Nuestro equipo ha analizado mediante electroforesis de campo pulsante (PFGE) diferentes cepas utilizadas en la elaboración de cavas, de las que se ha demostrado su inestabilidad genética. Usando una cepa silvestre aislada y caracterizada, DC5, hemos realizado un estudio de inestabilidad cariotípica en crecimiento vegetativo, analizando alrededor de 500 cariotipos. A partir de este cúmulo de datos hemos comprobado que esta cepa presenta una alta frecuencia de inestabilidad cariotípica, cercana a 10-2 cambios por generación. Aunque aparentemente esta cepa es excepcional en este sentido, existen cepas enológicas de levadura con tasas algo menores, pero también importantes, por lo que el control de este fenómeno es crucial para el seguimiento de las fermentaciones industriales a gran escala (figuras 1 y 2).
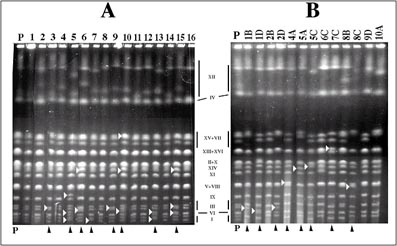
B) Cariotipos de esporas de DC5: El carril P muestra el cariotipo de DC5. Para cada espora, el número que aparece en su nombre se refiere al asca de la cual procede; las cuatro esporas individuales se denominan de A hasta D. Ninguna asca dio más de dos esporas viables. Las esporas que muestran reordenaciones cromosómicas están marcadas. Las asignaciones de cromosomas de las cepas se han hecho por similaridad electroforética con la cepa de laboratorio W303a (Nadal et al.,1999). Nótese las variaciones el cromosoma XII (marcadas con bandas verticales). Las técnicas genéticas de aislamiento de esporas han permitido obtener derivados estables a partir de cepas inestables
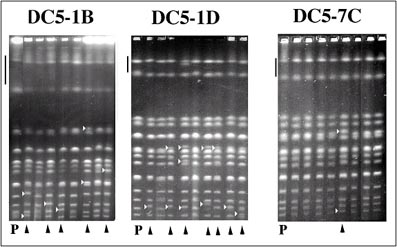
Una manera de abordar el problema de la inestabilidad cromosómica es a través del análisis genético del fenómeno de inestabilidad cariotípica. Las células de la cepa DC5 tienen la capacidad de esporular, es decir, producir esporas que heredan sus características genéticas siguiendo las normas de herencia mendeliana clásicas, aunque con algunas pequeñas peculiaridades derivadas del ciclo vital asexual de la levadura. Una sorpresa de este análisis fue que el 70% de estos derivados meióticos (que llamaremos derivados F1) presentan una menor inestabilidad cariotípica, tanto meiótica (inmediatamente tras esporular) como mitótica, del orden de 10-4 cambios por generación. Esta frecuencia es cercana a la de mutación espontánea y, por tanto, estos clones pueden ser considerados como clones genéticamente estables. Los derivados F1 pueden volver a esporular (reesporulación), y los derivados F2 resultantes muestran mayoritariamente una baja frecuencia de inestabilidad cromosómica durante el crecimiento vegetativo. Por tanto, esta es una manera de conseguir derivados genéticamente estables de una cepa inestable.
Otra manera de reducir la inestabilidad de una cepa es destruyendo los genes necesarios para que el fenómeno tenga lugar. Esto es lo que se hizo con DC5, a la que se eliminaron las copias del gen rad52, necesario para la recombinación homóloga y, por tanto, supuestamente responsable de gran parte de las reordenaciones cromosómicas. Los derivados deficientes en RAD52 (Drad52) demostraron en efecto ser más estables que la cepa parental, pero sin llegar a la estabilidad de algunos derivados F1 o F2. Por tanto, se puede deducir que el proceso de inestabilidad genética es sólo en parte dependiente de recombinación homóloga.
Pruebas de segunda fermentación
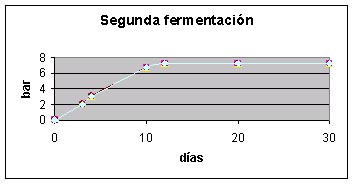
Se realizaron pruebas de segunda fermentación para asegurar la idoneidad enológica de las cepas estables obtenidas. La levadura se multiplicó a partir de medio sólido en mosto estéril y después se adaptó al crecimiento en el vino base con seguimiento de lecturas de turbidez. Una vez hecho el recuento de las poblaciones se inoculó a razón de 106 células viables por mL de vino base. El vino y los envases también eran estériles. Se colocaron manómetros para hacer el seguimiento de la presión hasta el final de la fermentación. El vino base era un coupage de la cosecha 2000 con las variedades chardonnay, macabeo y parellada, que mostraban unos valores analíticos habituales:10% etanol, 6 g de acidez total y 24 g de sacarosa añadida (Cantos, 1992). Las botellas utilizadas eran las clásicas para cava, de 750 mL (adaptadas para soportar una presión de hasta 10 bar). La presión se midió con manómetros adaptados a la botella con lecturas periódicas.
Como se observa en la figura 3, las cepas F1 de cariotipo estable fueron sometidas a pruebas fermentativas, junto con la cepa DC5, la DC5Drad52 y otras industriales. La fermentación de un vino base por parte de estas cepas fue igual a la de la cepa DC5 (inestable), manteniéndose las propiedades enológicas, en concreto la capacidad de fermentación vigorosa. Por tanto, es posible usar estos sistemas de estabilización del cariotipo para mejorar cepas industriales en las que la inestabilidad pueda representar un riesgo.
Cabe destacar que los procesos de esporulación y reesporulación son completamente naturales, equivalentes al establecimiento de líneas puras (razas) animales por retrocruzamiento y, por tanto, no presentan ningún conflicto con la legalidad vigente (y posiblemente futura). Además, nuestros resultados pueden contribuir a la comprensión de la inestabilidad del cariotipo en levaduras salvajes con potencial enológico e ilustran la posibilidad de remediación genética para aumentar la idoneidad de esas cepas para su uso industrial.
Conclusiones
Las levaduras utilizadas en la elaboración del cava pueden presentar una inestabilidad genética relativamente alta con frecuencias de entre 10-2 y 10-3, lo que puede comprometer su uso industrial. La esporulación de cepas inestables puede generar esporas estables con frecuencias de reordenaciones de 10-4 cambios por generación, frecuencia parecida a las cepas estables. La disrupción de los mecanismos de recombinación homóloga (cepas Drad52) también disminuye la inestabilidad cariotípica, pero sólo de manera parcial. Cualquiera que sea el método usado, las cepas estables poseen unas características fenotípicas óptimas para su utilización industrial. Así pues, presentaron las condiciones deseadas para este proceso industrial. Todas las fases de la elaboración (crecimiento en mosto, en vino y fermentación en botella) se produjeron sin los posibles problemas derivados de utilizar cepas inestables. Nuestros resultados contribuyen a la comprensión de la inestabilidad del cariotipo y ponen claramente de manifiesto la necesidad de realizar estudios de inestabilidad genética de las cepas industriales con el objeto de obtener derivados estables para su uso industrial.
Bibliografía
Bartra, E.: «Aspectos microbiológicos de la elaboración del cava», Microbiología SEM 1995 ; 11: 43-50.
Cantos, A.; Bartra, E.; Campamá, C. y Vilavella, M.: «Experiencias con levaduras secas comerciales en el tiraje del cava», Viticultura Enologia Profesional 1992; 22: 38-41.
Carro, D.; Piña, B.: Genetic analysis of the karyotype instability in natural wine yeast strains», Yeast 2001; 18 (16): 1457-1470.
Carro, D.; Piña, B.: «Identificació de soques de llevat de interés enològic», ACE Revista de Enologia 2000; 3: 10-14.
Carro, D.; García Martínez, J.; Pérez Ortín; Piña, B.: «Structural characterization of chromosome I size variants from a natural yeast strain», Yeast (en prensa).
Nadal, D.; Carro, D.; Fernández Larrea, J.B.; Piña B.: «Analysis and dynamics of the chromosomal complements of wild sparkling wine yeast strains», Appl Env Microbiol 1999; 65: 1688-1695.
Querol, A.; Barrio, E.; Ramón, D.: «A comparative study of different methods of yeast strain characterization», System Appl Microbiol 1992; 15: 439-446.

{kind=link}