

La obtención de vinos más atractivos y complejos desde el punto de vista organoléptico con el objetivo de satisfacer las necesidades del actual mercado global es uno de los objetivos prioritarios para los profesionales de la enología, que demandan nuevas y mejores cepas de levaduras adaptadas a diferentes tipos y estilos de vinos. Esta demanda puede satisfacerse a través de la selección de levaduras vínicas no Saccharomyces y su inclusión en cultivos iniciadores mixtos, capaces no solo de llevar a cabo la fermentación alcohólica sino de introducir características aromáticas diferenciales en los vinos obtenidos.
Durante la fermentación alcohólica las levaduras no sólo convierten los azúcares en etanol y dióxido de carbono, sino que también producen toda una serie de metabolitos volátiles que, aunque minoritarios, determinan de manera fundamental el carácter aromático del vino (figura 1). El perfil aromático de un producto tan complejo como el vino no es atribuible a un solo compuesto de impacto, sino que es el resultado de la combinación e interacción entre los distintos compuestos aromáticos. A pesar de ello, su aroma genérico de fondo se atribuye mayoritariamente a alcoholes y ésteres, que le otorgan su calidad e intensidad aromática.
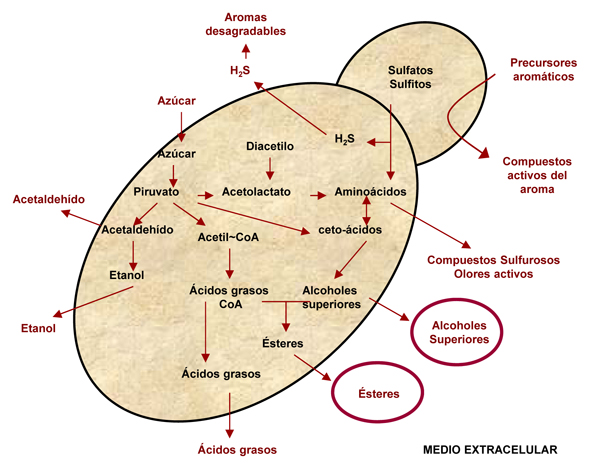
[Fuente: Tomada de Rojas 2002]
Los ésteres, compuestos volátiles con fragancias aromáticas agradables, son generados en pequeñas cantidades pero a una concentración superior a su umbral de percepción. Los más importantes son los ésteres polares, responsables de dar al vino cuerpo y consistencia, y los ésteres apolares, donde se incluyen los ésteres de acetato de alcoholes superiores y los ésteres de etilo de ácidos grasos saturados. En general, los ésteres apolares se asocian al aroma frutal, floral y fresco de los vinos jóvenes (tabla 1).
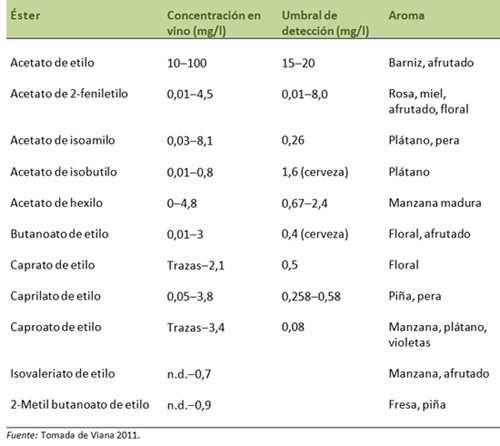
Diferentes estudios han documentado la capacidad de las levaduras no Saccharomyces para sintetizar ésteres de acetato. Por ejemplo, su capacidad para sintetizar acetato de etilo es, en general, superior a la de las cepas vínicas de Saccharomyces cerevisiae, y determinados géneros como Rhodotorula, Pichia y Hansenispora han sido señaladas como buenas productoras de acetato de isoamilo y acetato de 2-feniletilo. Además, estos estudios han demostrado que las levaduras no Saccharomyces pueden ser seleccionadas en base a su capacidad para producir metabolitos secundarios que contribuyan a mejorar la calidad del vino.
En este contexto, nuestra propuesta para conferir mayor complejidad aromática a los vinos ha sido la selección de levaduras no Saccharomyces que, formando parte de cultivos mixtos con S. cerevisiae, sean capaces de incrementar la concentración de acetato de 2-feniletilo, ya que este compuesto se asocia con una aroma afrutado, floral y con notas de miel y tabaco (Viana, 2011).
Escrutinio de cepas: importancia de los criterios de selección
Las levaduras no Saccharomyces se han asociado tradicionalmente a la formación de ésteres de acetato, sobre todo acetato de etilo, cuya incidencia negativa sobre la calidad del vino cuando se superan ciertos niveles (150-200 mg/L) llevó a catalogar a estas levaduras como alterantes. Sin embargo, la producción de estos compuestos es dependiente de cepa, por lo que unos criterios racionales de selección permitirían identificar aquellas levaduras más apropiadas.
Nuestra propuesta para un primer escrutinio en medio microbiológico fue emplear como criterio de selección la capacidad para combinar una alta producción de acetato de 2-feniletilo con bajos niveles de acetato de etilo. El escrutinio se realizó empleando 38 cepas de levaduras de origen vínico, pertenecientes a la Colección Española de Cultivos Tipo (CECT). Se incluyeron especies del género Candida (C. cantarelli, C. dattila y C. stellata), Hanseniaspora (H. guilliermondii, H. uvarum y H. vineae), Pichia (P. anomala, P. fermentans y P. membranifaciens), Torulaspora (T. delbrueckii) y Zygosaccharomyces (Z. bailii y Z. rouxii). A efectos comparativos, se incluyeron también en el estudio 9 cepas comerciales de S. cerevisiae. Como resultado de este escrutinio (figura 2), se seleccionaron 4 cepas de levadura pertenecientes a los géneros Hanseniaspora y Pichia (H. guilliermondii 11027 y 11102, H. vineae 1471, P. membranifaciens 10550).
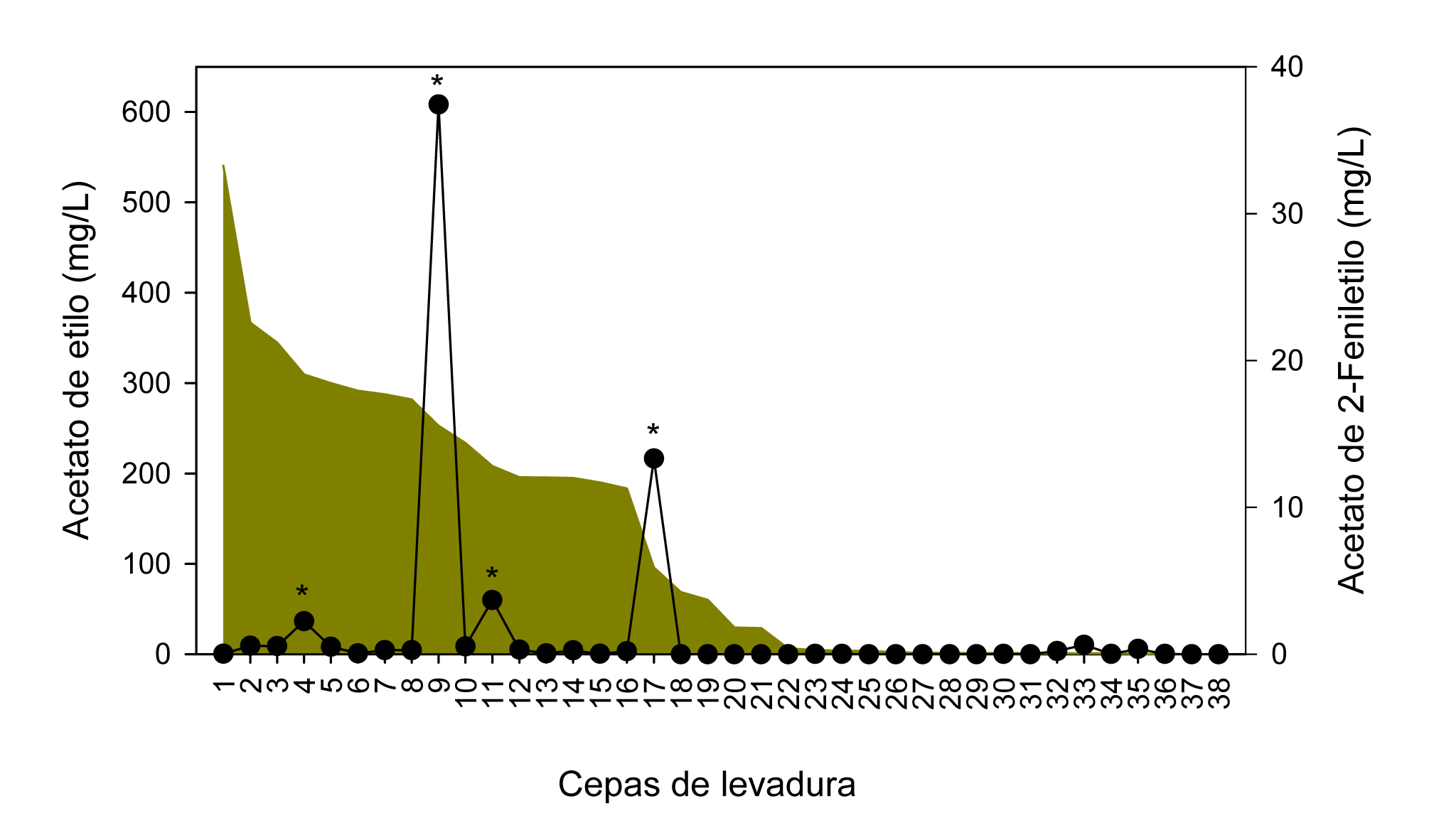
El segundo criterio de selección fue la baja producción de SH2 y de vinil- y etilfenoles a través de las actividades sulfito reductasa y cinamato descarboxilasa, dado que estos compuestos impactan negativamente sobre la calidad del vino. La actividad sulfito reductasa no se detectó en la cepa de P. membranifaciens, pero sí en las tres cepas de Hanseniaspora, aunque a niveles similares a los detectados en las cepas comerciales de S. cerevisiae. La actividad cinamato descarboxilasa no se detectó en ninguna de las cepas seleccionadas. Por lo tanto se siguió adelante con las 4 cepas para completar su caracterización.
« El ácido acético puede inhibir el crecimiento de S. cerevisiae, (…). Este efecto de inhibición también se asocia a concentraciones excesivas de ácidos grasos de cadena corta, capaces de provocar paradas de fermentación. »
Con el objetivo de seleccionar aquella cepa más apropiada para formar parte de un cultivo iniciador mixto con S. cerevisiae, el tercer criterio de selección fue el perfil enológico. Este perfil incluyó consumo de glucosa y fructosa, por su posible efecto sobre el comportamiento fermentativo de S. cerevisiae debido a su naturaleza glucofílica. Así mismo también se consideró la formación de etanol, acidez volátil, acetaldehído, glicerol, ácidos grasos de cadena corta, ésteres y alcoholes superiores. En concreto, el ácido acético, además de suponer un problema organoléptico cuando su concentración se aproxima a su umbral de detección (0,7-1,1 mg/L), también puede inhibir el crecimiento de S. cerevisiae, aunque para ello son necesarias concentraciones notablemente superiores (4,5-7,5 g/L). Este efecto de inhibición también se asocia a concentraciones excesivas de ácidos grasos de cadena corta, capaces de provocar paradas de fermentación.
De las levaduras evaluadas, P. membranifaciens 10550 fue descartada por su incapacidad de crecer en mosto. Las tres cepas restantes, en mayor o menor medida, fermentaron el mosto y produjeron niveles de acetaldehído, glicerol y ácido decanoico ligeramente inferiores a los niveles producidos por S. cerevisiae, mientras que los niveles de ácido hexanoico y octanoico y la concentración de alcoholes superiores fueron notablemente inferiores a S. cerevisiae, por lo que estos compuestos no se consideraron determinantes como criterio de exclusión. La producción de ácido acético se situó en el intervalo 0,57-1,09 g/L, correspondiendo el valor más alto a H. guilliermondii 11102, por lo que fue descartada. Finalmente, la formación de acetato de etilo y de 2-feniletilo nos llevó a seleccionar a H. vineae 1471 (acetato de etilo: 39,9 mg/L; acetato de 2-feniletilo: 12,9 mg/L) frente a H. guilliermondii 11027 (acetato de etilo: 114,8 mg/L; acetato de 2-feniletilo: 1,55 mg/L) para formar parte de un cultivo iniciador mixto.
Prueba de concepto en mosto estéril: diseño del cultivo iniciador mixto
Una vez seleccionada H. vineae 1471, se abordó el diseño de cultivos iniciadores mixtos. Para ello, se evaluaron distintos iniciadores que incluyeron como base la proporción entre no Saccharomyces y S. cerevisiae que es frecuente al inicio de las fermentaciones espontáneas (90:10, % H. vineae: % S. cerevisiae), así como cultivos en los que se fue reduciendo de forma progresiva el porcentaje de H. vineae (75:25, 50:50, 25:75, 10:90, 5:95 y 0:100). La finalidad de este objetivo fue el diseño de iniciadores más acordes con el punto de vista del bodeguero, que tiende a valorar mejor aquellos cultivos que contienen porcentajes elevados de S. cerevisiae.
A la vista de los resultados obtenidos (figura 3), fue posible reducir el porcentaje de H. vineae hasta un 50%, sin comprometer el efecto beneficioso buscado, ya que la cantidad de acetato de 2-feniletilo obtenida fue significativamente superior a la producida por S. cerevisiae.
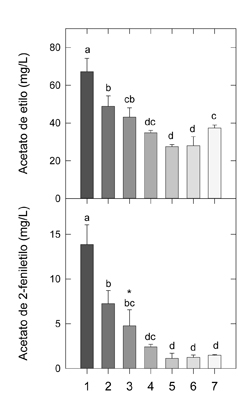
Otra conclusión extraída de estos experimentos fue que, independientemente de su porcentaje inicial, H. vineae fue capaz de crecer en presencia de S. cerevisiae de forma similar a su crecimiento en cultivo puro. En nuestras condiciones de fermentación, la población de H. vineae decreció tras alcanzar el máximo después de 2 días de fermentación, hasta su práctica desaparición a los 8 días, con la excepción de los vinos inoculados con el mayor porcentaje. Tradicionalmente, se ha asociado la desaparición de las levaduras no Saccharomyces, particularmente las especies del género Hanseniaspora, después de los primeros 3-4 días de fermentación con su poca tolerancia al etanol. Sin embargo, en concordancia con nuestros resultados, hay cada vez más estudios que demuestran que las levaduras apiculadas pueden sobrevivir a lo largo de la fermentación alcohólica durante períodos mucho más largos que los inicialmente descritos.
Prueba de concepto en mosto natural: influencia del tipo de inoculación
Teniendo en cuenta que las fermentaciones realizadas en bodegas no se realizan con mostos estériles sino con mostos que, a pesar de ser tratados con sulfuroso, presentan una microbiota propia, se estudió la implantación de algunos de los cultivos iniciadores diseñados en mostos naturales no estériles. Para ello se empleó mosto de la variedad Tempranillo, con un recuento inicial de levaduras elevado (>106 cfu/ml). El 90% de estas levaduras fueron no Saccharomyces y aunque no hubo ninguna especie claramente predominante, H. guilliermondii, H. osmophila y Zygosaccharomyces cidri, representaron, cada una de ellas, entre un 20 y un 30% de la microbiota inicial. Es de destacar que en el mosto no se detectó H. vineae, lo que facilitó el seguimiento por técnicas moleculares de la cepa inoculada (Viana et al., 2011a).
Dado que algunas firmas comerciales recomiendan la aplicación de los cultivos mixtos de forma secuencial para conseguir una mejor implantación, se incluyó este tipo de inoculación (primero se inocula la levadura no Saccharomyces y transcurridos varios días se inocula S. cerevisiae) comparándola con la inoculación conjunta (se inoculan ambas levaduras simultáneamente). El tipo de inoculación no influyó de manera notable sobre la cinética de fermentación, aunque sí sobre las especies predominantes. En la inoculación secuencial, H. vineae 1471 fue la especie de levadura no Saccharomyces predominante al inicio de la fermentación (días 1 y 3), lo que no se observó en el caso de la inoculación conjunta. En concordancia con el mayor crecimiento de la cepa de H. vineae en las fermentaciones secuenciales, la concentración de acetato de 2-feniletilo (1,7 mg/L) fue superior, con significancia estadística, a la obtenida en los vinos inoculados conjuntamente (0,8 mg/L). Para poner en contexto estas cantidades de acetato de 2-feniletilo con las cantidades más elevadas obtenidas en mostos estériles, hay que recordar que el porcentaje mínimo de H. vineae en el iniciador para que se alcanzara una concentración significativa del éster fue del 50%. Con este porcentaje se alcanzó una concentración celular de H. vineae superior a la de S. cerevisiae durante los tres primeros días de fermentación, lo que no fue posible en el caso del mosto no estéril a pesar de utilizar un inóculo 90:10. Es obvio que la presencia de la microbiota nativa en este último caso influyó en el crecimiento de H. vineae y por tanto no se alcanzaron concentraciones tan elevadas de acetato de 2-feniletilo como las obtenidas en mostos estériles. Por último, señalar que no se puede descartar la contribución de la microbiota nativa a la concentración final de acetato de 2-feniletilo, sobre todo teniendo en cuenta la presencia de H. guilliermondii en el mosto y durante los primeros días de fermentación, ya que se trata de una especie con reconocida capacidad de formación de dicho éster.
Inmovilización de H. vineae y su empleo en una inoculación mixta secuencial
Una estrategia efectiva para controlar el período de permanencia de las levaduras no Saccharomyces en el mosto en fermentación es la inmovilización celular. Esta estrategia se ha aplicado con éxito en el caso de Schizosaccharomyces pombe al inicio de la fermentación alcohólica para lograr el consumo parcial o total de ácido málico y evitar, una vez conseguido el efecto, el desarrollo de aromas extraños característicos de esta levadura.
«Una estrategia efectiva para controlar el período de permanencia de las levaduras no Saccharomyces en el mosto en fermentación es la inmovilización celular.»
En el caso concreto de H. vineae, la inmovilización, además de asegurar la práctica desaparición de la levadura, permitiría modular la cantidad de acetato de 2-feniletilo en función del momento de la retirada. Por ello, tras comprobar que el entrampamiento de H. vineae en esferas de alginato no afectaba a la capacidad de formación de acetato de 2-feniletilo, se seleccionaron 3 tiempos a los cuales retirar las células inmovilizadas e inocular S. cerevisiae en forma libre. Considerando que el período de permanencia se relaciona con la producción del éster, se seleccionaron los días 4, 5 y 6 de fermentación, correspondientes a un consumo del 25, 50 y 60 % de los azúcares iniciales. Tras la inoculación con S. cerevisiae se completó la fermentación, obteniéndose vinos con todos los parámetros enológicos dentro de los rangos considerados adecuados y lo que es más importante, con concentraciones diferentes de acetato de 2-feniletilo: 2,7 mg/L cuando se retiró H. vineae el cuarto día, 4,7 mg/L cuando se retiró el quinto, y 6,4 mg/L al retirar la levadura inmovilizada tras 6 días de fermentación (Viana et al., 2011b).
Consideraciones finales
Considerando todos los resultados expuestos, resulta evidente que la utilización de cultivos iniciadores mixtos con levaduras no Saccharomyces seleccionadas con criterios enológicos razonables es una alternativa prometedora que merece la pena continuar explorando.
Los resultados demuestran que H. vineae 1471, como integrante de cultivos iniciadores, produce vinos cuyos parámetros enológicos se encuentran en el intervalo descrito como apropiado, y es capaz de incrementar los niveles de acetato de 2-feniletilo con respecto a los producidos por S. cerevisiae. Sin embargo, dada la complejidad aromática del vino, habría que estudiar en mayor profundidad el impacto del incremento de un único compuesto sobre el aroma global. Es interesante destacar que bien a través de la proporción de H. vineae en el iniciador mixto como a través de su inmovilización, es posible modular la concentración de acetato de 2-feniletilo. Ambas estrategias son atractivas para el enólogo y resultaría imprescindible profundizar en su estudio.
Por último, un aspecto muy importante que no se ha abordado en estos estudios, y que facilitaría la aplicación de H. vineae a escala industrial, sería su producción en forma de levadura seca activa como los iniciadores de S. cerevisiae. Es de esperar que dada la experiencia de las empresas elaboradoras de levaduras, la producción de diferentes especies de no Saccharomyces en forma de LSA no presente grandes dificultades, y que en los próximos años se incremente la oferta comercial de este tipo de iniciadores.
Agradecimientos
Este trabajo fue financiado por el MINECO a través de los proyectos AGL2003-01295 y AGL2004-00978. Los autores agradecen a todos sus colaboradores por su contribución a este trabajo.
Bibliografía
. Rojas V.: Actividades esterásicas en levaduras vínicas. Tesis Doctoral, Universidad de Valencia, 2002
. Viana F.: Levaduras no Saccharomyces para modular el aroma secundario de los vinos: incremento del acetato de 2-feniletilo mediante cultivos iniciadores mixtos. Tesis Doctoral, Universidad Politécnica de Valencia, 2011.
. Viana F., Gil J. V., Genovés S., Vallés S., Manzanares P.: Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiology 2008; 25: 778-85.
. Viana F., Gil J. V., Vallés S., Manzanares P.: Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. International Journal of Food Microbiology 2009; 135: 68-74.
. Viana F., Belloch C., Vallés S., Manzanares P.: (2011a). Monitoring a mixed starter of Hanseniaspora vineae – Saccharomyces cerevisiae in natural must: impact on 2-phenylethyl acetate production. International Journal of Food Microbiology 2011a; 151: 235-40.
. Viana F., Taillandier P., Vallés S., Strehaiano P., Manzanares P.: 2-Phenylethyl acetate formation by immobilized cells of Hanseniaspora vineae in sequential mixed fermentations. American Journal of Enology and Viticulture 2011b; 62: 122-6.


{kind=link}