

Se está constatando el cambio climático con un progresivo aumento de las temperaturas en nuestras latitudes. Ello está conllevando a un aumento de azúcares en el mosto de uva, que da lugar a vinos con mayor grado alcohólico. Debido a esto, las bacterias lácticas como Oenococcus oeni que deben realizar la fermentación maloláctica tienen mayores dificultades para ello, que se añaden a las ya de por sí difíciles condiciones de pH bajo, pocos nutrientes, etc. La mayor maduración de la uva también comporta niveles de L-málico menores. Todo ello requiere un mejor conocimiento de los mecanismos de adaptación de O. oeni a estas condiciones estresantes. Las nuevas herramientas moleculares, como las técnicas de análisis global (transcriptómica, proteómica y metabolómica) aportan la posibilidad de tener una visión completa de la respuesta celular y de conocer cual es la importancia relativa de cada uno de los mecanismos de protección que intervienen. Ello contribuirá a proponer nuevos protocolos de adaptación de los cultivos iniciadores de fermentación maloláctica, para conseguir su correcto desarrollo en vinos difíciles para O. oeni.
Dificultades en el inicio de la fermentación maloláctica: vinos con elevado grado alcohólico y bajo contenido en ácido L-málico
La fermentación maloláctica (FML) sigue siendo objeto de estudio debido a las dificultades que puede presentar su inicio después de la fermentación alcohólica. Oenococcus oeni, especie mayoritariamente responsable de dicho proceso, ha sido descrita como la bacteria láctica más resistente a las condiciones estresantes del vino. Actualmente, el mercado ofrece una amplia oferta de cepas de O. oeni seleccionadas que pueden ser usadas como cultivos iniciadores para asegurar el desarrollo de la FML. A pesar de ello, en ocasiones, la FML puede ser incompleta o no tener lugar. Entre los múltiples factores causantes de las dificultades en el desarrollo de O. oeni, cabe destacar la pobreza de nutrientes en el vino y los factores inhibidores como el bajo pH y el elevado contenido en etanol. Este último, previsiblemente, tomará mayor relevancia debido al calentamiento global y el aumento del grado alcohólico en los vinos de regiones cálidas. De hecho, en ciertas zonas vitivinícolas, como la D.O.Q. Priorat o D.O. Montsant, los elevados grados alcohólicos que presentan ya actualmente sus vinos suponen a menudo una importante limitación para el desarrollo de la FML.
Existen numerosos trabajos sobre la respuesta al estrés etanólico. El principal efecto es la acidificación intracelular debido a la fluidización de la membrana. La disminución del pH interno provoca la inhibición de múltiples funciones celulares. Entre los mecanismos descritos como protectores frente a dicho estrés se encuentran la modificación de la composición tanto lipídica como proteica de la membrana citoplasmática (Silveira et al., 2004, Chu-Ky et al., 2004). De este modo, se ha propuesto como posible protocolo de preadaptación de cultivos iniciadores de O. oeni la aplicación de diversos choques (ácido, frío, etanol) para aclimatar las células antes de su inoculación en el vino (Chu-Ky et al., 2004).
Otro de los factores que presumiblemente afectará al desarrollo de O. oeni será el bajo contenido de ácido L-málico debido al cambio climático y el consecuente mayor grado de madurez de la uva. En algunos vinos de regiones cálidas, como las antes mencionadas, ya se están registrando actualmente valores de ácido L-málico alrededor de 0,5 g/L. Dada la pobreza en nutrientes del vino y los requisitos nutricionales de O. oeni (descrita como fastidiosa desde el punto de vista de su crecimiento), la poca disponibilidad de ácido L-málico, principal substrato utilizable por O. oeni en el vino, puede comprometer la supervivencia de esta bacteria láctica.
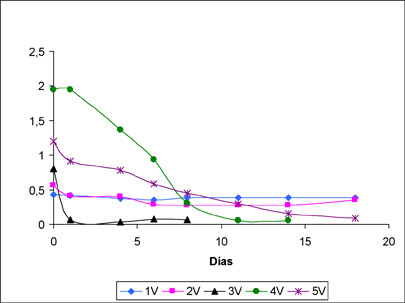
A modo de ejemplo, en la figura 1 se muestra el desarrollo de la FML en un ensayo con cinco vinos tintos diferentes inoculados con una misma cepa de O. oeni. Como se puede observar, la concentración inicial de ácido L-málico en los diferentes vinos, que oscila entre 2 y 0,5 g/L aproximadamente, influye en gran modo en el desarrollo de la FML. En los dos vinos con valores de ácido L-málico próximos a 0,5 g/L de ácido L-málico, 1V y 2V, no llega a desarrollarse la FML, mientras que en el resto de vinos se finaliza en todos los casos en un plazo de entre 2 y 18 días. Cabe destacar, que los dos vinos en los que no tuvo lugar la FML tenían un contenido en etanol diferente. Mientras que el vino 1V contenía un 14,5% de etanol (v/v), el vino 2V contenía un 12 % de etanol. Así pues, estos datos indicarían el carácter limitante de una baja concentración de ácido L-málico, independientemente de otros factores inhibidores, como el propio etanol. Todo ello apunta a la necesidad de estudiar cómo O. oeni puede afrontar la poca disponibilidad de su principal sustrato en el vino. Por otra parte, se abre la cuestión de si las cepas seleccionadas por su resistencia a otros factores de estrés funcionarán con la misma eficiencia ante la falta de ácido L-málico y de si podrían ser preadaptadas en este sentido.
La utilización del ácido cítrico ayuda a O. oeni a sobrevivir en el vino
En carencia de otros sustratos, como el ácido L-málico, el ácido cítrico puede ser un nutriente alternativo para O. oeni. El consumo de este ácido orgánico, presente en el vino en concentraciones variables, normalmente cercanas a 0,5 g/L, puede modificar el perfil organoléptico del vino. Por cada molécula de ácido cítrico se genera como mínimo una de ácido acético, incrementando de forma ineludible la acidez volátil. Por otra parte, la vía metabólica de consumo de ácido cítrico puede llevar a la producción de compuestos aromáticos como el diacetilo, aportando el carácter de “mantequilla” al vino. Dicho carácter puede resultar más o menos deseable en función del estilo de vino que se quiera producir (Bartowsky y Henschke, 2004). A pesar de que en la mayoría de cepas de O. oeni el consumo de ácido cítrico se inicia hacia el final de la FML (Bartowsky y Henschke, 2000), en vinos con un bajo contenido inicial de ácido L-málico, la utilización del ácido cítrico se puede dar de forma casi simultanea al desarrollo de la FML, lo que puede suponer un riesgo para aquellos vinos en los que no se desee la aparición de diacetilo.
En contrapartida, la utilización de citrato resulta beneficiosa para el desarrollo de O. oeni ya que le aporta energía a la célula en forma de ATP. De hecho, en estudios recientes se ha podido relacionar el metabolismo del ácido cítrico con la respuesta al estrés etanólico (Olguín et al., 2009). En un ensayo realizado en vino sintético se pudo demostrar que el consumo de ácido cítrico en relación a la biomasa incrementa notablemente por efecto de la presencia del 10% de etanol (tabla 1).

Además, se pudo observar una mayor producción de ácido acético por mol de ácido cítrico consumido, lo que indicaría que, en presencia de etanol, el flujo metabólico de la vía del ácido cítrico se desviaría hacia la conversión del piruvato en acetato, con la consiguiente formación de ATP. De todo ello se deduce que O. oeni utiliza el ácido cítrico para hacer frente al estrés, consiguiendo energía adicional y, probablemente, contrarrestando la acidificación interna como se ha descrito para otras bacterias lácticas, como Lactococcus lactis, debido a la eliminación de protones del citoplasma en el proceso de metabolización del citrato (Martín et al., 2004).
En este mismo estudio se pudo comprobar la activación de la expresión de genes como citE, que codifica el enzima citrato liasa (responsable de convertir el ácido cítrico en oxalacetato y ácido acético), por la presencia de etanol a bajo pH. Los genes, que como el mencionado muestran una respuesta transcripcional al estrés asociado al vino, podrían ser utilizados como marcadores para la selección de cepas, como indicadores de mayor resistencia y, en el caso de citE, también de consumo más activo de ácido cítrico.
Perspectivas: estudio molecular global y búsqueda de nuevos protocolos de adaptación celular
Más allá de los estudios individuales de diversos mecanismos de respuesta al estrés en O. oeni, las nuevas herramientas moleculares, como las técnicas de análisis global (transcriptómica, proteómica y metabolómica), permitirán el estudio en mayor profundidad de O. oeni y su resistencia a las condiciones adversas. Estas técnicas aportan la posibilidad de tener una visión completa de la respuesta celular y de conocer cuál es la importancia relativa de cada uno de los mecanismos de protección que intervienen. Hasta el momento, sólo han sido publicados trabajos basados en la aplicación de técnicas proteómicas al estudio de O. oeni.
El análisis del proteoma (conjunto de todas las proteínas que conforman la célula), con la aplicación de técnicas como la electroforesis en dos dimensiones (2-DE), permite visualizar el conjunto de proteínas celulares y cuantificar los cambios en su concentración en función de las condiciones a la que se exponen los cultivos. La proteómica presenta todavía algunas limitaciones técnicas en cuanto al poder de detección (no se llegan a detectar todas las proteínas) y la posterior identificación de las proteínas (por falta de secuencias publicadas en las bases de datos de referencia). En este sentido, la figura 2 muestra un gel 2-DE que presenta una buena resolución, en base a lo que permiten las técnicas actualmente en uso, del proteoma citosólico de O. oeni. En dicho gel se pueden ver un total de 215 spots, donde cada uno se asociaría a una proteína diferente.

El primer trabajo sobre O. oeni a nivel proteómico fue el de Silveira et al. en 2004, y posteriormente en 2009 se publicó el estudio de Cecconi et al. Ambos trabajos abordan el estudio del efecto del etanol sobre O. oeni, comparando células adaptadas con células no adaptadas. A pesar de las diferencias en las condiciones de ensayo, en ambos estudios se pudo detectar el claro incremento de la concentración de enzima glutatión reductasa en células adaptadas al etanol. Este resultado indicaría la importancia del mantenimiento del balance redox celular en la adaptación al estrés. Por otra parte, Silveira et al. (2004), además de las proteínas citosólicas estudiaron también las proteínas de membrana y asociadas a membrana. De dicho estudio se pudo observar el notable incremento de proteínas asociadas a membrana en células adaptadas al etanol.
Estos primeros resultados, apuntan a algunos de los mecanismos que serían más relevantes, en el global de la célula, en la lucha contra el estrés de O. oeni. Todo ello no es más que una pincelada del potencial de las técnicas de análisis global, como la proteómica, que permiten analizar los elementos celulares (proteínas, genes, metabolitos, etc.) de forma completa y conjunta.
De todos modos, queda mucho camino por recorrer en el campo de las “ómicas” en lo que se refiere a O. oeni. Para avanzar en el conocimiento de esta peculiar bacteria, todavía queda por estudiar su respuesta y adaptación al estrés a nivel transcriptómico (expresión de los genes) y metabolómico (cambios en los compuestos consumidos o producidos en las diferentes vías metabólicas), así como profundizar y mejorar su estudio proteómico. Todo ello ha de contribuir a la propuesta de nuevos protocolos de adaptación de cultivos iniciadores de FML, aspecto clave, mas allá de la selección de cepas, para conseguir el correcto desarrollo de la FML en vinos difíciles para O. oeni.
Agradecimientos
Este trabajo se enmarca en los proyectos financiados Cenit-Deméter del CDTI (Plan Ingenio 2010) y AGL2009-07369 del Plan Nacional.
Bibliografía
Bartowsky, E.J., Henschke, P.A. 2000. Management of malolactic fermentation for the ‘buttery’ diacetyl flavour in wine. Australian Grapegrower and Winemaker 438a, 58–67.
Bartowsky, E.J., Henschke, P.A. 2004. The ´buttery` attribute of wine – diacetyl – desirability, spoilage and beyond. International Journal of Food Microbiology 96, 235– 252.
Cecconi, D., Milli, A., Rinalducci, S., Zolla, L., Zapparoli, G. 2009. Proteomic analysis of Oenococcus oeni freeze-dried culture to assess the importance of cell acclimation to conduct malolactic fermentation in wine. Electrophoresis, 30: 2988-2995.
Chu-Ky, S., Tourdot-Marechal, R., Marechal, P.-A., Guzzo, J. 2005. Combined cold, acid, ethanol shocks in Oenococcus oeni: Effects on membrane fluidity and cell viability. Biochimica et Biophysica Acta 1717, 118-124.
Martín, M.G., Sender, P.D., Peirú, S., de Mendoza, D., Magni, C. 2004. Acid-inducible transcription of the operon encoding the citrate lyase complex of Lactococcus lactis Biovar diacetylactis CRL264. Journal of Bacteriology 186, 5649-5660.
Olguín, N., Bordons, A., Reguant, C. 2009. Influence of ethanol and pH on the gene expresión of the citrate pathway in Oenococcus oeni. Food Microbiology 26, 197-203.
Olguín, N. 2010. Tesis Doctoral: Molecular study of the mechanisms of Oenococcus oeni involved in tis adaptation to wine conditions and in the development of malolactic fermentation. Universitat Rovira i Virgili.
Silveira, M.G., Baumgärtner, M., Rombouts, F.M., Abee, T. 2004. Effect of adaptation to ethanol on cytoplasmic and membrane protein profiles of Oenococcus oeni. Applied and Environmental Microbiology 70, 2748-2755.



{kind=link}